Forum geobotanicum (2006) 2: 24-44
DOI 10.3264/FG.2006.1013
Reprint (PDF) Version
(click here) ca. 4 MB
Daniela Röder*, Michael Jeschke, Kathrin Kiehl
Lehrstuhl für Vegetationsökologie, Technische Universität München
Vegetation und Böden alter und junger Kalkmagerrasen im Naturschutzgebiet „Garchinger Heide“ im Norden von München
Vegetation and soils of ancient and young calcareous grasslands in the nature reserve “Garchinger Heide” (
Published online: 13 October 2006
© Forum geobotanicum 2006
Abstract The nature reserve „Garchinger Heide“ north of Munich (Bavaria, Germany) is a remnant of the formerly widespread calcareous grasslands of the Munich Gravel Plain on dry, nutrient-poor calcaric Regosol soils with low water-storage capacity. Most of the nature reserve is covered by species-rich ancient grasslands with a high proportion of species of the Mesobromion erecti and Cirsio-Brachypodion. At the “Rollfeld”, where topsoil was removed in 1945, vegetation cover is lower and the proportion of species of the Xerobromion, Sedo-Scleranthetea and Seslerietalia albicantis is higher than on the ancient grasslands due to low water-availability and low nutrient contents. In 2003, the number of ancient grassland species, which have successfully established on the topsoil-removal site, was higher than in vegetation descriptions from 1956 and 1986. On an ex-arable field within the nature reserve, which was last ploughed in 1959, several species of the Molinio-Arrhenatheretea were still found in 2003. In comparison to 1986, however, the number of species of the Mesobromion erecti and Cirsio-Brachypodion was higher and the vegetation has become more similar to the ancient grassland vegetation. This positive development was also reflected in the soil analyses; the contents of exchangeable P2O5 and K2O in the soil of the ex-arable field have decreased considerably during the last 12 years.
Keywords calcareous grassland, topsoil removal, ex-arable field, vegetation, soil nutrients
Zusammenfassung
Das Naturschutzgebiet „Garchinger Heide“ ist ein Relikt der ehemals ausgedehnten Kalkmagerrasen auf Pararendzinen über Niederterrassenschotter im Alpenvorland. Bei jahrhundertelanger extensiver Landnutzung hat sich auf trockenen nährstoffarmen Böden eine artenreiche Vegetation mit zahlreichen seltenen und gefährdeten Arten entwickelt. Die „Altheide“, welche den größten Teil des Naturschutzgebiets einnimmt, ist geprägt durch flachgründige, nährstoffarme Böden, welche eine geschlossene Vegetation mit einem hohen Anteil an Arten der Halbtrockenrasen (hauptsächlich Mesobromion erecti und Cirsio-Brachypodion) tragen. Das 1945 durch Oberbodenabtrag begonnene, aber nicht mehr ausgebaute und verwendete „Rollfeld“ weist einen niedrigen Feinbodenanteil, welcher auf eine geringe Wasserverfügbarkeit schließen lässt, sowie niedrige Gehalte an Gesamtstickstoff und CAL-austauschbaren P2O5 auf. Die Vegetation ist lückig mit einem hohen Anteil an Trockenrasenarten (Xerobromion, Sedo-Scleranthetea, Sesleritalia albicantis). Das heutige Vorkommen zahlreicher Arten, welche in früheren Untersuchungen (1956 und 1986) nicht gefunden wurden, lässt auf eine fortschreitende Sukzession der Vegetation des Rollfeldes hin zu der der Altheide schließen. Der 1959 zum Naturschutzgebiet hinzu gekaufte ehemalige Acker unterscheidet sich immer noch deutlich von der Altheide. Eine allmähliche Annäherung der Standortbedingungen ist jedoch zu beobachten. So sanken die Gehalte an CAL-austauschbarem P2O5 und K2O im Vergleich zu Werten aus dem Jahr 1993 deutlich ab. Trotz des immer noch hohen Anteils an Grünlandarten (Molinio-Arrhenatheretea) konnten sich im Vergleich zu 1986 mehr Magerrasenarten etablieren.
-------------------------------------------------
* Korrespondierende Autorin:
Daniela Röder
Am Hochanger 6, 85350 Freising
Email: roederd@wzw.tum.de
Tel.: +49 8161 - 714142
Fax: +49 8161 - 714143
-------------------------------------------------
Einleitung
Mitteleuropäische Kalkmagerrasen, die auch als Grasheiden bezeichnet werden, gehören zu den Vegetationstypen mit der höchsten kleinräumigen Artenvielfalt (alpha-Diversität) der Welt (Peet et al. 1983, Willems et al. 1993). Durch die Änderung der Landnutzung im 20. Jahrhundert ging jedoch ein Großteil der ehemals ausgedehnten Magerrasen verloren (Willems 2001, Pfadenhauer 2002, Poschlod und WallisDeVries 2002). Die heutigen Kalkmagerrasenreste sind stark fragmentiert und liegen isoliert in einer zumeist stark genutzten Agrarlandschaft (Quinger et al. 1994; Ringler 2002). Die Kalkmagerrasen der Mittelgebirge (z.B. Fränkische und Schwäbische Alb) wurden überwiegend aufgeforstet bzw. der Sukzession überlassen (Beinlich und Manderbach 1995, Bender et al. 2005). Ein Teil der Flächen am Rand von Ortschaften ist heute bebaut (Bender et al. 2005). Die meisten Kalkmagerrasen auf den würmeiszeitlichen Schmelzwasserschottern des Voralpenlandes am Lech oder an der Isar wurden mit Hilfe moderner Geräte und Düngemethoden in Ackerland umgewandelt oder im Zuge des Kiesabbaus zerstört (Vollmann 1911, Riemenschneider 1956). Im Naturschutzgebiet „Garchinger Heide“ konnten 27 Hektar der ehemals ausgedehnten Kalkmagerrasen im Norden von München durch die Initiative der Bayerischen Botanischen Gesellschaft zu Beginn des 20. Jahrhunderts vor dem Umbruch in Ackerland gerettet werden.
Die Vegetation der „Garchinger Heide“ besteht aus subkontinentalen, ursprünglich beweideten Magerrasen mit submediterranen, dealpinen und pontischen Florenelementen (Vollmann 1911, Liebermann 1986). Aufgrund dieser Kombination stuft Kaufhold (1990) diese Grasheiden als international bedeutsam ein. Sie sind nach Art. 20c Bundesnaturschutzgesetz und Art. 13d Bayerisches Naturschutzgesetz geschützte Standorte (Bayerisches Landesamt für Umweltschutz 2000), gehören zum Lebensraumtyp 6210 gemäß Anhang I der FFH-Richtlinie (Lang et al. 2004) und gelten als stark gefährdet nach der Roten Liste der Pflanzengesellschaften Deutschlands (Rennwald 2000). Mehr als 50 der insgesamt 218 nachgewiesenen Blütenpflanzen stehen auf der Roten Liste der gefährdeten Farn- und Blütenpflanzen Bayerns (Scheuerer und Ahlmer 2003). Mit Pulsatilla patens (L.) Mill. ist auch eine Pflanzenart des Anhangs II der FFH-Richtlinie vertreten (Council of Europe 1979). Nach Korneck et al. (1993) stellt die Magerrasenvegetation der „Garchinger Heide“ eine Art Vorposten der in Osteuropa weit verbreiteten subkontinentalen Steppen des Cirsio-Brachypodion (Klasse Festuco-Brometea) mit der Assoziation Adonido-Brachypodietum pinnati dar.
In der „Garchinger Heide“ wurden aufgrund ihres Artenreichtums und des Vorkommens zahlreicher seltener Pflanzen- und Tierarten bereits ab Mitte des 19. Jahrhunderts wissenschaftliche Untersuchungen durchgeführt (z.B. Sendtner 1854, Vollmann 1911, Riemenschneider 1956, Hepp und Poelt 1970, Pfadenhauer und Liebermann 1986, Korneck et al. 1993, Miller und Pfadenhauer 1997, Kiehl und Jeschke 2005, Jeschke und Kiehl 2006). Im Verlauf des 20. Jahrhunderts kam es im jetzigen Naturschutzgebiet immer wieder zu Veränderungen, wie z.B. zum Abtrag des Oberbodens in einzelnen Teilbereichen, zur Erweiterung durch den Ankauf angrenzender Äcker oder zu Änderungen des Pflegeregimes. Damit ging auch ein Wandel der Vegetationszusammensetzung und -struktur sowie der Bodeneigenschaften einher.
Ziel dieser Untersuchung ist daher, verschiedene Standorte des Naturschutzgebietes mit Hilfe von Bodendaten und Vegetationsaufnahmen genauer zu charakterisieren und die Beziehung zwischen Vegetation und Standorteigenschaften zu klären. Dabei sollen nicht nur die Veränderungen in den letzten 50 Jahren diskutiert werden, sondern auch Perspektiven für die zukünftige Entwicklung des Naturschutzgebiets aufgezeigt werden.
Untersuchungsgebiet
Lage, Geologie und Klima
Das 27 ha große Naturschutzgebiet „Garchinger Heide“ liegt in der Münchner Schotterebene etwa 15 km nördlich von München an der Landstraße von Eching nach Dietersheim (48°18' N, 11°39' E, 469 m über NN) (Abb. 1). Die Umgebung des Naturschutzgebietes ist geprägt durch intensive Ackernutzung sowie große Siedlungs- und Verkehrsflächen. Westlich der „Garchinger Heide“ verläuft die stark befahrene Autobahn A9 München-Berlin.
Das Naturschutzgebiet „Garchinger Heide“ ist Teil der naturräumlichen Einheit „Münchner Schotterebene“ (Wittmann 1983). Die Ebene ist die größte würmeiszeitliche Schotterfläche (Niederterrassenschotter) nördlich der Alpen (Pfadenhauer 2002) und dehnt sich zwischen den Endmoränen des ehemaligen Isargletschers und dem Tertiärhügelland bzw. dessen südlich vor gelagerten Grundwassermooren aus (Fetzer et al. 1986, Windolf 1989).
Das Klima im Bereich des Naturschutzgebiets ist schwach subkontinental mit einem sommerlichen Niederschlagsmaximum. Der durchschnittliche Jahresniederschlag liegt bei 883 mm und die Jahresmitteltemperatur bei 9 °C mit einem mittleren Minimum im Januar von -2°C und einem mittleren Maximum im Juli von 18°C (Deutscher Wetterdienst, Wetterstation Oberschleißheim, langjährige Mittel 1961-1990).

Abb. 1 Lage des Naturschutzgebiets „Garchinger Heide“ nördlich von München (Bayern, BRD, 48°18' N, 11°39' E, 469 m NN). Die Streifen im südlichen Teil des Gebiets sind durch die kurz vorher durchgeführte Streifenmahd entstanden. Quelle: Bodeninformationssystem Bayern, www.bis.bayern.de, Befliegung 2003.
Fig. 1 Location of the nature reserve „Garchinger Heide“north of Munich (Bavaria, Germany, 48°18' N, 11°39' E, 469 m NN). The stripes in the south of the reserve result from the mowing shortly before. Source: Bodeninformationssystem Bayern, www.bis.bayern.de, Flight 2003.
Geschichte der Landnutzung und aktuelles ManagementDie Besiedlung der „Garchinger Heide“ kann bis in die Bronzezeit (1800 – 1000 vor Chr.) zurückverfolgt werden (Kollmansberger und Geisel 1989a). Aus dieser Zeit stammen zwei Hügelgräbergruppen im Nordwesten und Südwesten des Naturschutzgebiets. Im frühen Mittelalter legten die Bajuwaren im Gebiet Hochäcker an, um Ackerbau auf den kargen Böden zu betreiben (Kollmansberger und Geisel 1989b). Reste solcher Hochäcker können im Osten des Naturschutzgebiets gefunden werden. Bis zum Ende des 19. Jahrhunderts war jedoch ein Großteil der Schotterebene allgemeines Weideland (Allmende) für Schafe, Rinder, Ziegen und Schweine oder einschürige Mähwiese (Sendtner 1854). Durch diese Nutzung entstand eine weite, durch niedrigwüchsige Kalkmagerrasen geprägte Landschaft, die nur vereinzelt von Gehölzinseln durchbrochen wurde (Pfadenhauer 2002). Noch 1850 erstreckten sich diese Grasheiden im Münchner Norden über eine Fläche von schätzungsweise 15.000 ha (Sendtner 1854, Pfadenhauer 2001). Ende des 19. Jahrhunderts wurde die Allmende unter den Hofbesitzern der Gegend aufgeteilt. Dies schuf die rechtliche Grundlage für die folgende Intensivierung der landwirtschaftlichen Nutzung. Um einen Teil der artenreichen Kalkmagerrasen vor der Umwandlung in Ackerland zu bewahren, kaufte die Bayerische Botanische Gesellschaft zwischen 1907 und 1914 insgesamt 23 ha Land (Geisel 1989). Im Jahr 1942 wurde diese Fläche offiziell als Naturschutzgebiet „Garchinger Heide“ ausgewiesen (Kollmannsberger und Geisel 1989c). Aufgrund weiterer Flächenankäufe in den Jahren 1933 und 1959 umfasst das Gebiet heute 27,13 ha (Geisel 1989).
Als Pflegenutzung wird im Naturschutzgebiet „Garchinger Heide“ eine Streifenmahd durchgeführt. Dabei wird die Zentralfläche in Streifen von ca. 40 m Breite eingeteilt, die bis 1998 jährlich alternierend im Oktober gemäht wurden, seit 1999 für die Mähgutübertragung früher (Ende Juli bis September). Für jeden einzelnen Streifen entsprach dies einer Mahd im zweijährigen Turnus. Um der Verfilzung der Vegetation und der Zunahme der Streuschicht entgegenzuwirken, wurde seit 2004 der „Bracheanteil“ reduziert, so dass nun jährlich wechselnd jeweils drei Streifen gemäht werden und nur der vierte Streifen „brach“ liegt. Die Randbereiche und der ehemalige Acker werden jährlich Ende Juli/Anfang August gemäht. Das Mähgut wird auf ehemaligen Ackerflächen für die Wiederansiedlung von Magerrasen verwendet (Pfadenhauer und Miller 2000, Pfadenhauer und Kiehl 2003, Kiehl et al. 2006).
Das Naturschutzgebiet „Garchinger Heide“ stellt ein wichtiges Naherholungsziel für die Bewohner der Umgebung dar. So ergibt sich besonders zum Blühzeitpunkt der meisten Heidearten im Frühjahr und Sommer ein starker Besucherdruck. Um diesen gezielt zu lenken, wurde ein Wegekonzept mit zwei Hauptwegen und wenigen kleinen Nebenwegen eingeführt. Trotzdem werden immer wieder seltene Pflanzenarten durch Betreten der Flächen und Sammeln beeinträchtigt.
Methoden
Untersuchungsstandorte
Ziel der bodenkundlichen und vegetationskundlichen Untersuchungen war, die drei wichtigsten Standorttypen des Naturschutzgebiets „Garchinger Heide“ zu charakterisieren. Der größte Teil des Gebiets wird von der Altheide eingenommen, d.h. von ursprünglichen, nie umgebrochenen Grasheiden (Abb. 2). Die Mächtigkeit des humusreichen feinkörnigen Oberbodens erreicht hier im Durchschnitt nur 10-20 cm (Pfadenhauer und Liebermann 1986). Im südlichen Teil der Altheide liegt das 1,6 ha große Rollfeld (Abb. 2). In den letzten Kriegsmonaten sollte hier im Jahr 1945 durch Häftlinge aus dem Konzentrationslager Dachau auf einer Breite von durchschnittlich 40 m und einer Länge von etwa 300 bis 400 m durch Abtragen des Oberbodens per Hand eine Landebahn für den Militärflughafen Schleißheim geschaffen werden (Kollmannsberger und Geisel 1989d). Wegen des Kriegsendes wurde das Rollfeld jedoch nie fertig gestellt und benutzt . Bis heute konnte sich hier nur ein 1 - 2 cm mächtiger Ah-Horizont über dem anstehenden Kies bilden. Somit stellt das Rollfeld einen Standort für anspruchslose, konkurrenzschwache Gefäßpflanzen sowie zahlreiche Moos- und Flechtenarten dar (Jeschke und Kiehl 2006).
Im Jahr 1959 wurde ein ca. 1,6 ha großer Acker im Osten des Naturschutzgebiets hinzugekauft (Abb. 2). Bei dem Boden dieses ehemaligen Ackers handelt es sich um eine Ackerpararendzina. Im Laufe der Jahre konnten sich hier auch ohne künstliche Ansiedlungsversuche zahlreiche Kalkmagerrasenarten etablieren; außerdem kommen aber auch einige anspruchsvolle Arten des Wirtschaftsgrünlandes (Klasse Molinio-Arrhenatheretea) vor, die ansonsten im Naturschutzgebiet fehlen (Pfadenhauer und Miller 2000, Kiehl und Jeschke 2005).

Abb. 2 Standorttypen im Naturschutzgebiet „Garchinger Heide“. Altheide: ursprüngliche Magerrasen, Rollfeld: Bodenabtrag 1945, ehemaliger Acker: bis 1959 bewirtschaftet.
Fig. 2 Habitat types of the nature reserve “Garchinger Heide“. Ancient grassland, “Rollfeld” (topsoil removal 1945) and ex-arable field (last ploughed in 1959).
Bodenkundliche UntersuchungenZur Beschreibung der Böden des Naturschutzgebiets stellte das Bayerische Geologische Landesamt bisher unveröffentlichte Bodenprofile und dazu gehörige Analysedaten aus dem Jahr 1985 zur Verfügung (Lage siehe Abb. 2). Die Bezeichnung der Horizonte richtet sich nach der Bodenkundlichen Kartieranleitung (Ad-Hoc-Arbeitsgruppe Boden 2005). Die Anlage von aktuellen Bodenprofilen war aufgrund der mit dem Eingriff verbundenen großen Störung des Naturschutzgebietes nicht möglich. Im September 2004 wurden im Bereich der Altheide sowie auf dem Rollfeld jeweils 15 Bodenproben und auf dem ehemaligen Acker fünf Proben des Oberbodens (0-10 cm) entnommen. Aufgrund des sehr hohen Skelettgehalts des Bodens konnte keine volumengerechte Beprobung mittels Stechzylinder durchgeführt werden. Die Bodenproben wurden bei 60°C getrocknet, gewogen und gesiebt (2 mm Maschenweite). Der Feinboden (< 2 mm) wurde gewogen und hinsichtlich des pH-Werts und der Nährstoffgehalte analysiert. Jeweils die Hälfte einer Bodenprobe wurde gemahlen, um mit Hilfe eines Elementaranalysators (VarioEL) im CN-Modus den Gehalt an Gesamtstickstoff und Gesamtkohlenstoff zu bestimmen. In kalkhaltigen Böden tritt Kohlenstoff nicht nur in organischer (Corg) sondern auch in anorganischer Form (Canorg) als Bestandteil des Kalziumcarbonats (CaCO3) auf. Deshalb wurde der Kalziumcarbonatanteil der Bodenproben nach der Scheibler-Methode (Schlichting et al. 1995) ermittelt und daraus der Anteil des anorganischen Kohlenstoffes berechnet. Die Differenz des Gesamtkohlenstoffs und des anorganischen Kohlenstoffs ergab dann den Gehalt an organischem Kohlenstoff, der für die Berechnung des C/N-Verhältnises verwendet wurde. Um das Angebot an pflanzenverfügbarem Phosphor und Kalium in den Bodenproben abzuschätzen, wurde der Gehalt an Kalziumlactat-(CAL-)austauschbarem P2O5 und K2O bestimmt (Schlichting et al. 1995, VDLUFA 2002). Der pH-Wert wurde in CaCl2-Lösung gemessen (VDLUFA 1991).
Vegetationskundliche UntersuchungenUm die Vegetation der „Garchinger Heide“ zu beschreiben, wurden in den Jahren 2003 und 2004 insgesamt 104 Vegetationsaufnahmen auf 4 m² großen Flächen durchgeführt. Diese Flächengröße wurde gewählt, um eine Vergleichbarkeit mit Angaben anderer Untersuchungen in Kalkmagerrasen der Münchner Schotterebene (z.B. Kiehl und Wagner 2006, Will et al. 2005) sowie weiteren europäischen Kalkmagerrasen (z.B. Dengler 2005) zu gewährleisten. Es wurden 62 Flächen im Bereich der Altheide aufgenommen, 26 Flächen auf dem Rollfeld und 16 Flächen auf dem ehemaligen Acker. Die Deckung aller auf den Aufnahmeflächen vorkommenden Gefäßpflanzenarten wurde in Prozent geschätzt. Außerdem wurden die Gesamtdeckung der Gefäßpflanzen, Moose und Strauchflechten, der Streu sowie die Deckung des unbedeckten Bodens (inklusive Steine) ermittelt. Die auf dem Rollfeld vorkommenden Krustenflechten (vgl. Schauer 1969) wurden in der vorliegenden Arbeit nicht untersucht. Die Nomenklatur der Gefäßpflanzen richtet sich nach Oberdorfer (2001, 8. Auflage).
DatenauswertungFür die Gefäßpflanzen wurde die Artenzahl pro 4 m²-Fläche ermittelt sowie der Anteil magerrasentypischer Arten und Grünlandarten an der Artenzahl pro 4 m² bestimmt. Die magerrasentypischen Arten umfassten die beiden folgenden Gruppen: Arten der Halbtrockenrasen und Arten der Trockenrasen. Zu den Arten der Halbtrockenrasen gehören vor allem Arten der Verbände Mesobromion erecti Br.-Bl. et Moor 1938 und Cirsio-Brachypodion Had. et Klika 1944 (nach Oberdorfer 2001). Auch einzelne Arten der Klassen Erico-Pinetea Horvat 1959 und Trifolio-Geranietea sanguinei Th. Müll. 1961, die typisch für genutzte Kalkmagerrasen im Münchner Norden sind, wurden dieser Gruppe zugeordnet. Arten des Verbands Xerobromion Br.-Bl. et Moor 1938 sowie der Klassen Sedo-Scleranthetea Br.-Bl. 1955 em. Th. Müll. 1961 und Seslerietalia albicantis Br.-Bl. 1948 wurden der Gruppe „Arten der Trockenrasen“ zugeordnet. Gefäßpflanzenarten des Wirtschaftsgrünlands (Klasse Molinio-Arrhenatheretea Tx 1937) bilden die dritte Gruppe „Grünlandarten“. Für alle Vegetationsaufnahmen wurden für die Seggen als Matrixbildner, die als konkurrenzstark geltenden Süßgräser, die stickstofffixierenden Leguminosen, die Chamaephyten, welche besonders typisch für nie umgebrochenen Trocken- und Halbtrockenrasen sind und für sonstige Krautarten, die prozentualen Anteile an der Artenzahl sowie an der Deckungssumme (Summe aller prozentual geschätzten Deckungswerte) bestimmt. Dabei wurden die nur in wenigen Aufnahmen vorkommenden Gehölze (Salix eleagnos Scop., Rhamnus saxatilis Jacq.) mit zu den sonstigen Krautigen gezählt, da eine gesonderte Darstellung dieser im Gebiet niedrigwüchsigen Gehölze aufgrund der geringen Anzahl nicht sinnvoll war. Um eine doppelte Zuordnung von Arten zu vermeiden wurden der Gruppe Chamaephyten nur diejenigen Arten zugeordnet, die nicht zur Familie der Fabaceae gehören. Auf die Ergebnisse hat dies keinen Einfluss, da die chamaephytischen Leguminosen auf allen Standorten mit ähnlichen Artenzahlen und Deckungen vertreten waren. Weiterhin wurden für alle Standorte die Matrixarten, d.h. die Arten, die in mindestens 90% der Aufnahmeflächen eines Standorts mit einer durchschnittlichen Deckung von mehr als 5% vorkommen, ermittelt.
Um Vegetationsgradienten zu beschreiben, wurde mit den wurzeltransformierten Deckungswerten der Gefäßpflanzen eine DCA (Detrended Correspondence Analysis) berechnet. Arten, die in weniger als fünf Aufnahmen vorkamen (5%), wurden dabei ausgeschlossen. Der Anteil der durch die DCA erklärten Varianz wurde mit Hilfe einer "after-the-fact evaluation" unter Verwendung des Distanzmaßes der relativen Euklidischen Distanz bestimmt (McCune und Mefford 1999). Für die Interpretation der DCA wurden Korrelationen zwischen den „site scores“ der DCA und den Anteilen der Arten der Halbtrockenrasen, Trockenrasen und des Grünlands an der Artenzahl berechnet und auf Signifikanz geprüft. Dazu wurde der Spearman-Rangkorrelationskoeffizient rs verwendet. Für die Berechnung der Korrelation zwischen der Deckung von Brachypodium rupestre und der Anzahl der Magerrasen- bzw. Trockenrasenarten wurde ebenfalls der Spearman-Rangkorrelationskoeffizient verwendet.
Unterschiede zwischen den Standorten hinsichtlich der Artenzahl, des prozentualen Anteils der oben genannten Artengruppen an der Artenzahl und der Deckung der Gefäßpflanzen, Moose, Strauchflechten, Streu, des offenen Bodens sowie der Chamaephyten, Leguminosen, Süßgräser, Seggen und sonstigen Krautigen wurden mit Hilfe des nichtparametrischen Mann-Whitney U-Tests paarweise analysiert. Auf die gleiche Weise wurden Unterschiede hinsichtlich des Feinbodenanteils, pH-Werts, der Gehalte an CAL-austauschbarem P2O5 und K2O sowie des Corg, des Nges und C/N-Verhältnisses getestet. Anschließend wurde eine Benjamini-Hochberg Korrektur für multiple Vergleiche durchgeführt (Benjamini und Hochberg 1995, Verhoeven et al. 2005).
Ergebnisse
Bodenkundliche Untersuchungen
Bei den Böden der „Garchinger Heide“ handelt es sich um Pararendzinen über Niederterrassenschotter (Abbildung 3). Der Ah-Horizont des in der Altheide aufgenommenen Profils besteht aus lehmigem Sand mit einem Kiesanteil von 10 bis 30 %. Der Lehmanteil des Feinbodens nimmt nach unten immer mehr ab bis im C-Horizont nur noch reiner Sand und Kies auftreten, wobei der kiesige Grobboden überwiegt. Der Humusgehalt nimmt im Bodenprofil vom Ah-Horizont zum C-Horizont ab, während der Carbonatgehalt ansteigt.

Abb. 3 Bodenprofil im Bereich der Altheide des Naturschutzgebiets „Garchinger Heide“ (Lage siehe Abb. 2). Mit freundlicher Genehmigung des Bayerischen Geologischen Landesamtes (www.geologie.bayern.de). Zusätze zu Horizontbezeichnungen: e=mergelig, l=Lockersubstrat, v=verwittert (nach Ad-Hoc-Arbeitsgruppe Boden 2005).
Fig. 3 Soil profile from the “Garchinger Heide“ ancient grassland in 1985 courtesy of Bayerisches Geologisches Landesamt (www.geologie.bayern.de). Soil depth, soil type (fine/coarse), % gravel, texture, organic content class, carbonate class (headings left to right). For location see Fig. 2. Additions within the description of the soil horizons: e=marly, l=unconsolidated, v=weathered (Ad-Hoc-Arbeitsgruppe Boden 2005).
Alle drei Standorte des Naturschutzgebiets unterscheiden sich hinsichtlich des Anteils an Feinboden, des Gehalts an CAL-austauschbarem K2O und des Gehalts an Gesamtstickstoff im Oberboden voneinander (Tabelle 1, Abb. 4). Der Gehalt an CAL-austauschbarem P2O5 ist an allen drei Standorten ähnlich. Der pH-Wert ist auf dem Rollfeld am niedrigsten und das C/N-Verhältnis auf dem ehemaligen Acker.
|
pH-Wert |
P2O5 |
K2O |
Nges [%] |
C/N-Verhältnis |
|
|
Rollfeld |
7,4 ± 0,1 a |
1,3 ± 0,8 a |
7,8 ± 1,4 a |
0,52 ± 0,09 a |
12,4 ± 0,5 a |
|
Altheide |
6,4 ± 0,3 b |
1,6 ± 1 a |
5,1 ± 1,3 b |
0,60 ± 0,09 b |
12,1 ± 0,3 a |
|
ehemaliger Acker |
7,4 ± 0,1 a |
1,6 ± 0,9 a |
11,2 ± 1,8 c |
0,68 ± 0,04 c |
11,5 ± 0,3 b |
Tab. 1: Ergebnisse der Analyse der bodenchemischen Analysen (pH-Wert, CAL-austauschbares P2O5 und K2O sowie C/N-Verhältnis). Angegeben sind die Mittelwerte ± Standardabweichung (Altheide n=15, Rollfeld n=15, ehemaliger Acker n=5). Der Feinbodenanteil bezieht sich auf den trockenen Boden. Alle übrigen Angaben beziehen sich auf den trockenen Feinboden (< 2 mm). Unterschiedliche Buchstaben bedeuten signifikante Unterschiede p < 0,05 (für jeden Parameter getrennt zu lesen).
Tab. 1: Results of the soil analyses: pH-value, CAL-exchangeable P2O5 and K2O and C/N-ratio. Values represent means ± standard deviation (ancient grassland n=15, “Rollfeld” n=15, ex-arable field n=5). The proportion of fine soil refers to soil dry weight. The other results apply to dry fine soil < 2 mm. Different letters show significant differences, p < 0.05 (to be read separately for each parameter).

Abb. 4 Mittlerer Anteil des Feinbodens und des organischen Kohlenstoffs im Boden. Dargestellt sind jeweils Mittelwert + 1 Standardabweichung. Unterschiedliche Buchstaben bedeuten signifikante Unterschiede, p < 0,05. Die unterschiedlichen Skalierungen sind zu beachten.
Fig. 4 Mean proportion of fine soil and percent of organic carbon (from left to right). Data represent mean values and + 1 standard deviation. Different letters show significant differences, p < 0.05. Note the different scales.
Alle drei Standorte des Naturschutzgebiets unterscheiden sich hinsichtlich des Anteils an Feinboden, des Gehalts an CAL-austauschbarem K2O und des Gehalts an Gesamtstickstoff im Oberboden voneinander (Tabelle 1, Abb. 4). Der Gehalt an CAL-austauschbarem P2O5 ist an allen drei Standorten ähnlich. Der pH-Wert ist auf dem Rollfeld am niedrigsten und das C/N-Verhältnis auf dem ehemaligen Acker.
Vegetationskundliche Untersuchungen
Die Ergebnisse der einzelnen Vegetationsaufnahmen sind in der Vegetationstabelle, Anhang 1, zu finden.
Deckung und Anteile verschiedener Artengruppen
Die Altheide und der ehemalige Acker sind sich in Bezug auf die Deckung der Gefäßpflanzen, Moose und Strauchflechten sowie der offenen Bodenstellen sehr ähnlich. Nur die Deckung der Streuschicht ist in der Altheide höher (Abb. 5). Auf dem Rollfeld ist die Deckung der Gefäßpflanzen und Moose deutlich geringer als an den anderen beiden Standorten. Die Deckung der Strauchflechten und der offenen Bodenstellen ist dagegen deutlich höher.
Die mittlere Artenzahl der Gefäßpflanzen pro 4 m²-Fläche unterscheidet sich nicht signifikant zwischen den drei Standorten. Auf dem Rollfeld treten im Durchschnitt 26 Gefäßpflanzenarten auf und in der Altheide sowie auf dem ehemaligen Acker 28. Den größten Anteil an der Artenzahl nehmen auf allen Standorten die Arten der Halbtrockenrasen ein (Abb. 6). Auf dem Rollfeld wurde der höchste Anteil der Arten der Trockenrasen gefunden und der niedrigste Anteil an Grünlandarten. Auf dem ehemaligen Acker verhielt es sich genau umgekehrt (Abb. 6). Die Anzahl der Magerrasen- und Trockenrasenarten korreliert negativ mit der Deckung von Brachypodium rupestre (Host) Roem. & Schult. (rs = -0,44, P < 0,05) (Abb. 7).
Auf dem Rollfeld waren der Anteil der Chamaephyten an der Artenzahl und Deckung sowie der Deckungsanteil der Seggen deutlich höher als an den andern beiden Standorten (Abb. 8). Der Anteil der Seggen an der Artenzahl war dagegen an allen drei Standorten ähnlich. Auf dem ehemaligen Acker war der Anteil der Süßgräser an der Artenzahl und Deckung signifikant am höchsten, während in der Altheide der Anteil der sonstigen krautigen Arten höher als an den anderen Standorten war.
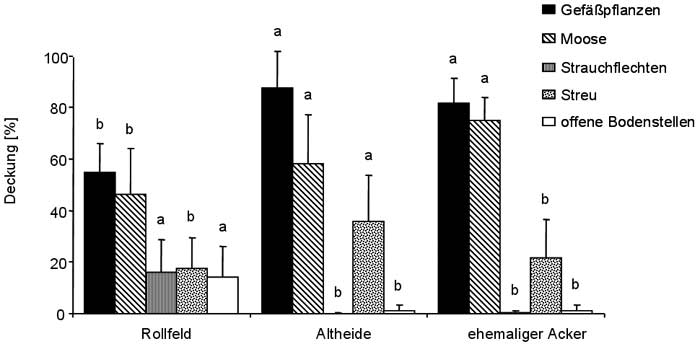
Abb. 5 Mittlere prozentuale Deckung der Gefäßpflanzen, Moose, Strauchflechten, Streu und der offenen Bodenstellen pro 4 m². Dargestellt sind jeweils Mittelwert + 1 Standardabweichung. Unterschiedliche Buchstaben bedeuten signifikante Unterschiede, p < 0,05 (für jeden Parameter getrennt zu lesen).
Fig. 5 Mean percentage cover of vascular plants, mosses, lichens, litter and open soil per 4 m² (from left to right). Data represent mean values + 1 standard deviation. Different letters show significant differences, p < 0.05 (to be read separately for each parameter).

Abb. 6 Mittlerer Anteil der Arten der Halbtrockenrasen (Mesobromion), Trockenrasen (Xerobromion) und des Wirtschaftsgrünlands (Molinio-Arrhenatheretea) an der Artenzahl auf 4 m². Unterschiedliche Buchstaben bedeuten signifikante Unterschiede, p < 0,05 (für jeden Parameter getrennt zu lesen).
Fig. 6 Mean proportion of species of the Mesobromion erecti and Cirsio-Brachypodion (black), species of the Xerobromion, Sedo-Scleranthetea and Seslerietalia albicantis (hatched) and Molinio-Arrhenatheretea species (white) in relation to species richness per 4 m². Different letters show significant differences, p < 0.05 (to be read separately for each parameter).

Abb. 7 Zusammenhang zwischen der Deckung von Brachypodium rupestre und der Anzahl der Magerrasen- und Trockenrasenarten in der Altheide n=62, Spearman-Rangkorrelationkoeffizient rs=-0,44, p<0,05.
Fig. 7 Correlation between the cover of Brachypodium rupestre and the number of dry grassland species (Festuco-Brometea, Sedo-Scleranthetea and Seslerietalia albicantis) in the ancient grassland n=62, rs = -0.44, p < 0.05.

Abb. 8 Mittlerer Anteil an der Artenzahl und Deckungsanteil der Leguminosen, Chamaephyten, Süßgräser, Seggen und sonstigen Krautigen. Unterschiedliche Buchstaben bedeuten signifikante Unterschiede, p < 0,05 (für jeden Parameter getrennt zu lesen).
Fig. 8 Mean proportion of legumes, chamaephytes, grasses, sedges and others herbaceous species in relation to species richness (left) and total cover (right). Different letters show significant differences, p < 0.05 (to be read separately for each parameter).
Analyse der Artenzusammensetzung
Die Ergebnisse der DCA (Detrended Correspondence Analysis) zeigen, dass sich die drei Standorte des Naturschutzgebietes (Rollfeld, Altheide, ehemaliger Acker) floristisch deutlich voneinander unterscheiden (Abb. 9). Entlang der ersten Achse, die 20,6% der Gesamtvarianz im Datensatz erklärt, wird der ehemalige Acker von der Altheide und dem Rollfeld getrennt. Auf der zweiten Achse, die weitere 14,2% der Varianz erklärt, werden das Rollfeld und die Altheide voneinander abgegrenzt. Der Anteil der Arten der Halbtrockenrasen korreliert negativ mit Achse 2 der DCA (rs=-0,59, P<0,001), während der Anteil der Trockenrasenarten negativ mit Achse 1 (rs=-0,46, P<0,001) und positiv mit Achse 2 (rs=0,48, P<0,001) korreliert. Der Anteil der Grünlandarten korreliert schwach positiv mit Achse 1 der DCA (rs=0,23, P=0,02).

Abb. 9 Ergebnis der DCA (Detrended Correpondence Analysis) mit den wurzeltransformierten Artmächtigkeiten. Die roten Vektoren stellen die Korrelationen des Anteils der Arten der Halbtrockenrasen, Trockenrasen und des Grünlands an der Artenzahl mit Achse 1 und 2 der Ordination dar. Die vollständigen Artnamen finden sich im Anhang 2.
Fig. 9 DCA-Ordination of the square-root transformed species cover data. The red vectors indicate correlations of the proportion of species of the Mesobromion erecti and Cirsio-Brachypodion; species of the Xerobromion, Sedo-Scleranthetea and Sesleritalia albicantis; and Molinio-Arrhenatheretea species with axis 1 and axis 2 of the DCA. For full names of plant species see appendix 2.
Als Matrixarten, die in mindestens 90% der Aufnahmeflächen mit einer durchschnittlichen Deckung > 5% vorkommen, wurden für die Altheide Carex humilis Leyss. und Anthericum ramosum L. ermittelt (Tabelle 2). Auf dem Rollfeld tritt lediglich Carex humilis als Matrixart auf. Außerdem kommen dort in fast allen Aufnahmeflächen Teucrium montanum L., Dorycnium germanicum (Gremli) Rikli und Thymus praecox subsp. praecox vor. Die dealpine Art Gentiana clusii Perr. & Song. ist ausschließlich auf das Rollfeld beschränkt. Der ehemalige Acker ist geprägt durch die Matrixarten Bromus erectus Huds. und Carex humilis. Dort kommt außerdem in allen Aufnahmeflächen Peucedanum oreoselinum (L.) Moench vor.
Folgende typische Magerrasenarten der Altheide fehlen in den untersuchten Flächen des ehemaligen Ackers: Adonis vernalis L., Carduus defloratus L. subsp. defloratus, Dorycnium germanicum (Gremli) Rikli, Erica herbacea L., Euphorbia verrucosa L., Euphrasia rostkoviana Hayne, Galium boreale L., Globularia cordifolia L., Inula hirta L., Polygala chamaebuxus L., Polygala amarella Crantz, Polygala vulgaris L., Polygonatum odoratum (Mill.) Druce, Potentilla alba L., Teucrium montanum L. Dagegen kommen einige andere Arten ausschließlich auf dem ehemaligen Acker vor: Achillea millefolium L. subsp. millefolium, Plantago lanceolata L., Ranunculus bulbosus L., Silene vulgaris (Moench) Garcke subsp. vulgaris, Taraxacum officinale agg.

Abb. 10 Altheide (Juli 2004); Foto: Daniela Röder
Fig. 10 Ancient grassalnd (July 2004); Photo: Daniela Röder

Abb. 11 Rollfeld (Mai 2001); Foto: Annuschka Thormann
Fig. 11 „Rollfeld“ (May 2001); Photo: Annuschka Thormann

Abb. 12 Ehemaliger Acker (Juli 2004); Foto: Daniela Röder
Fig. 12 Ex-arable field (July 2004); Photo: Daniela Röder
|
Rollfeld |
Altheide |
ehemalige Acker |
||||
|
Häufigkeit |
Deckung |
Häufigkeit |
Deckung |
Häufigkeit |
Deckung |
|
|
Anthericum ramosum |
38 |
0,8 |
100 |
27,0 |
81 |
2,0 |
|
Brachypodium rupestre |
65 |
1,0 |
82 |
9,3 |
69 |
6,6 |
|
Bromus erectus |
69 |
3,4 |
56 |
2,8 |
100 |
16,0 |
|
Carex humilis |
96 |
25,0 |
98 |
32,0 |
100 |
13,0 |
Tab. 2 Häufigkeit und mittlere Deckung von Anthericum ramosum, Brachypodium rupestre, Bromus erectus und Carex humilis auf dem Rollfeld, in der Altheide und auf dem ehemaligen Acker. Die Matrixarten, die in mind. 90% der Aufnahmeflächen mit einer durchschnittlichen Deckung > 5% vorkommen sind jeweils fett markiert.
Tab. 2 Frequency and mean cover of Anthericum ramosum, Brachypodium rupestre, Bromus erectus and Carex humilis at the “Rollfeld”, in the ancient grassland and on the ex-arable field. Matrix-species, which occur in at least 90% of the plots with an average cover of > 5%, are marked in bold.
Diskussion
Vergleich der Bodeneigenschaften an den untersuchten Standorten
Wasserverfügbarkeit und pH-Wert
Pararendzinen aus Schotter haben aufgrund ihres hohen Steingehalts eine geringe Wasserspeicherkapazität (Scheffer/Schachtschabel 2002). Tendenziell kann dabei davon ausgegangen werden, dass mit dem Feinbodengehalt des Bodens auch die Wasserverfügbarkeit steigt. Eine Studie von Leuschner (1989) zeigte unter Berücksichtigung des unterschiedlichen Skelettgehalts der Böden, dass die Wasserspeicherkapazität mit dem „physiognomischen Charakter“ der ihnen zugehörigen Rasengesellschaften korrespondiert und von Xerobromion über Mesobromion hin zu Arrhenatherion zunimmt. Demnach kann das Rollfeld auf Grund seines geringen Feinbodenanteils und des durch den Oberbodenabschub stark reduzierten Anteils organischen Kohlenstoffs als der Standort mit der geringsten Wasserverfügbarkeit im Naturschutzgebiet „Garchinger Heide“ bezeichnet werden. Der in der Altheide wesentlich höhere Anteil des Feinbodens und auch des organischen Kohlenstoffs im Oberboden lässt auf eine deutlich bessere Wasserverfügbarkeit schließen. Aufgrund des im C-Horizont anstehenden Schotters kommt es jedoch auch hier in niederschlagsarmen Perioden häufig zu Wasserknappheit. Der im Gegensatz zur Altheide geringere Feinbodenanteil des ehemaligen Ackers ist auf das frühere Pflügen der Fläche zurückführen, wodurch immer wieder größere Steine in den Oberboden gelangten und somit der Feinbodenanteil gesenkt wurde. Die Wasserverfügbarkeit ist deshalb auf dem ehemaligen Acker vermutlich etwas schlechter als in der Altheide.
Mit dem Anteil des Feinbodens und dem Verwitterungsgrad der Böden hängt auch der pH-Wert der Standorte eng zusammen. So war der pH-Wert auf dem Rollfeld und auf dem ehemaligen Acker auf Grund des niedrigeren Feinbodenanteils und damit höheren Skelettanteils kalkhaltigen Gesteins höher als in der Altheide. Zudem bewirkt in der Altheide auch der höhere Humusgehalt eine Absenkung des pH-Werts. Ein Vergleich der pH-Werte des Ah-Horizonts der Altheide zeigt in den letzten 35 Jahren nur leicht schwankende Werte um den pH-Wert 6 (Daten des Bayerisches Geologisches Landesamts von 1970 und 1987, Kiehl et al. 2003).
Nährstoffdynamik
Untersuchungen zu Aushagerung durch Mahd auf sandigen Böden in den Niederlanden zeigten, dass die Gehalte an austauschbarem Kalium schneller zurückgingen als die Phosphat-Gehalte des Bodens (Pegtel et al. 1996). Nach der Einstellung der Düngung und einer anhaltenden regelmäßigen Mahd kam es dort aber ebenso wie in einer Langzeitstudie in England und Wales (Skinner und Todd 1998) im Verlauf von 25 Jahren zu einem deutlichen Rückgang der Phosphat- und Kalium-Gehalte. Auch in der „Garchinger Heide“ lässt sich eine zunehmende Aushagerung durch Mahd feststellen. Vergleicht man die 2004 ermittelten Werte an CAL-austauschbaren P2O5 auf dem ehemaligen Acker mit Werten von Miller und Pfadenhauer (1997) aus dem Jahr 1993, so zeigt sich ein deutlicher Rückgang von Werten um 4,6 mg/100 g (1993) auf 1,6 mg/100 g (2004), was dem Niveau der Altheide im selben Jahr entspricht. Auch der Gehalt an Cal-austauschbarem K2O sank, jedoch nicht so schnell, von 20 mg/100 g im Jahr 1993 (Miller und Pfadenhauer 1997) auf 11,2 mg/100 g im Jahr 2004. Seit dem Ankauf des ehemaligen Ackers im Jahr 1959 konnten sich somit die Phosphat-Gehalte dem Niveau der Altheide angleichen und die Kalium-Gehalte annähern. Der Rückgang der P- und K-Gehalte in den Böden des ehemaligen Ackers der „Garchinger Heide“ deutet darauf hin, dass es inzwischen durch die jährliche Mahd ohne Düngung zu einer deutlichen Aushagerung des Bodens gekommen ist. Die Gesamtstickstoffgehalte auf dem ehemaligen Acker waren jedoch höher als im übrigen Untersuchungsgebiet, was vermutlich auf die höhere Deckung der Leguminosen, welche Stickstoff im Boden anreichern, zurückzuführen ist.
Untersuchungen auf Renaturierungsflächen mit Bodenabtrag im Umfeld der Garchinger Heide zeigten, dass bei einer durch Mähgutauftrag erhöhten Gesamtdeckung der Vegetation mehr Kalium im Oberboden angereichert wurde als auf Flächen mit geringer Vegetationsdeckung (Kiehl et al. 2003). So kann eventuell die Zunahme des Gehalts an CAL-austauschbarem K2O auf dem Rollfeld um 2,3 mg/100 g im Vergleich zum Jahr 2000 (Kiehl et al. 2003) mit der zunehmenden Gesamtdeckung der Vegetation durch die voranschreitende Sukzession erklärt werden. Möglicherweise spiegeln diese Werte jedoch auch nur kleinräumige oder zeitliche Schwankungen der Kalium-Verfügbarkeit wieder (vgl. Scheffer/Schachtschabel 2002). Die geringe Phytomasseproduktion auf dem Rollfeld (s. Kiehl et al. 2003) ist neben der Trockenheit vermutlich vor allem auf die geringe Stickstoffverfügbarkeit zurückzuführen, welche aus der geringen Wasserverfügbarkeit und dem geringen Gesamtstickstoffgehalt resultiert (Leuschner 1989, Neitzke 1998, Unkovich et al. 1998)
Zusammenfassend zeigen diese Ergebnisse, dass die Bedingungen für das Pflanzenwachstum vom Rollfeld über die Altheide hin zum ehemaligen Acker zunehmend günstiger werden.
Vegetationsentwicklung
Die drei Standorte des Naturschutzgebiets „Garchinger Heide” konnten auch nach einer Sukzession von 60 Jahren (Rollfeld) bzw. 45 Jahren (ehemaliger Acker) noch klar floristisch voneinander unterschieden werden. Die Einzigartigkeit der Altheide macht auch heute noch, wie bereits von Vollmann (1911) geschildert, das gleichzeitige Vorkommen submediterraner (z.B. Asperula cynanchica L., Hippocrepis comosa L.), dealpiner (z.B. Gentiana clusii, Globularia cordifolia) und pontischer (z.B. Adonis vernalis, Pulsatilla patens) Florenelemente aus. Die von Riemenschneider (1956) beschriebene Artenzusammensetzung scheint sich aber verändert zu haben. So dominieren heute Carex humilis und Anthericum ramosum sowie Brachypodium rupestre und Bromus erectus. Koeleria pyramidata (Lam.) P. Beauv. und Koeleria macrantha (Ledeb.) Schult. wurde im Gegensatz zu 1956 im Jahr 2004 in den Vegetationsaufnahmen der Altheide nicht gefunden. Carex montana L. und Avenula pratensis (L.) Dumort. traten nur in einzelnen Aufnahmeflächen mit geringen Deckungen auf. Besonders Brachypodium rupestre hat jedoch, wie auch die nah verwandte Art Brachypodium pinnatum (L.) P. Beauv., (During und Willems 1984, Bobbink und Willems 1987, Hurst und John 1999) einen negativen Einfluss auf die Artenzahl und in unserer Untersuchung besonders auf die Anzahl der Magerrasen- und Trockenrasenarten. In der Altheide wurde Brachypodium rupestre vermutlich einerseits durch die zu extensive Mahdnutzung im zweijährigen Turnus gefördert und zusätzlich durch den Eintrag von Stickstoff aus der Luft durch Verkehr- und Industrie-Emissionen (vgl. Bobbink und Willems 1987, Carroll et al. 2003, Jacquemyn et al. 2003).
Aufgrund der hohen Deckung offener Bodenstellen und des geringen Feinbodenanteils konnten sich auf dem Rollfeld besonders konkurrenzschwache Arten der Trockenrasen ansiedeln, die an den anderen Standorten zum Teil durch höher wüchsige Arten verdrängt werden. Riemenschneider (1956) beobachtete auf dem Rollfeld noch Initialstadien der Magerrasen mit geringen Deckungen von Thymus praecox, Leontodon incanus (L.) Schrank und Anthyllis vulneraria subsp. alpestris (Kit. ex Schult.) Asch. & Graebn. Pfadenhauer und Liebermann (1986) beschrieben einen Rückgang dieser Pionierarten und eine Zunahme von Süßgräsern und Carex humilis. Heute ist Carex humilis die Matrixart des Rollfeldes. Auch weitere typische Arten der Altheide konnten sich hier ausbreiten, wie z.B. Pulsatilla patens (Röder und Kiehl 2006). Die gesamte Artenzusammensetzung des Standortes gleicht heute der, welche Riemenschneider (1956) für kiesige Böden mit lehmig-feinsandigem Material und lückigem Vegetationsschluss beschrieben hat, mit Vorkommen von Carex humilis, Teucrium montanum, Thesium linophyllon L., Potentilla incana P. Gaertn., B. Mey. & Scherb. und Scabiosa canescens Waldst. & Kit. Trotz dieser fortschreitenden Sukzession ist das Rollfeld heute noch sehr heterogen. So wechseln Standorte mit fast geschlossener Vegetationsdecke kleinräumig mit Bereichen mit sehr lückiger Vegetation. Dadurch ergibt sich ein Mosaik, das momentan noch vielen konkurrenzschwachen Arten einen Lebensraum bietet (vgl. Jeschke und Kiehl 2006). Bei weiter zunehmender Deckung der Gefäßpflanzen und Moose könnte es aber künftig zum Rückgang von Arten kommen, die auf einen hohen Anteil offener Bodenstellen angewiesen sind. Für solche Pflanzenarten und vor allem für die für das Rollfeld typischen Tierarten wurde 1980 und 2000 auf zwei an das Rollfeld angrenzenden Flächen der Oberboden abgeschoben, deren Vegetationsentwicklung bei Will et al. (2005) beschrieben wird.
Nicht nur in der Altheide und auf dem Rollfeld, sondern auch auf dem ehemaligen Acker kommt heute eine hohe Anzahl von Arten der Halbtrockenrasen vor. Aufgrund des geringen Anteils der Trockenrasenarten ist der Anteil typischer Magerrasenarten insgesamt jedoch wesentlich niedriger als an den anderen beiden Standorten. Auch Untersuchungen in England zeigen, dass sich Magerrasen auf ehemaligen Äckern nur sehr langsam der Vegetationszusammensetzung alter Grasländer annähern. Dort konnten auch nach 100 Jahren noch Unterschiede zwischen der Vegetation ehemaliger Äcker und ursprünglicher Kalkmagerrasen festgestellt werden (Wells et al. 1976, Gibson und Brown 1991). Der Hauptgrund für diese Unterschiede ist vermutlich die langsame Ausbreitung der Zielarten (Hutchings und Booth 1996). Carex humilis, die von Pfadenhauer und Liebermann (1986) als fehlend auf dem ehemaligen Acker beschrieben wurde, tritt heute bereits in fast allen Aufnahmeflächen auf. Da Carex humilis durch Ameisen ausgebreitet wird (Oberdorfer 2001), verlief die Ausbreitung offenbar schneller als bei anemochoren Arten, wie z.B. Pulsatilla patens (vgl. Röder und Kiehl 2006). Auch eine erfolglose Keimung oder Etablierung konkurrenzschwacher Arten im unmittelbaren Umfeld konkurrenzstarker Gräser und Grünlandarten sowie eine dichte Moosschicht könnte die Ansiedlung von Magerrasen- und Trockenrasenarten auf dem ehemaligen Acker erschweren. Wie bereits 1986 beschrieben (Pfadenhauer und Liebermann 1986), konnte auch heute noch eine Reihe typischer Grünlandarten auf dem ehemaligen Acker gefunden werden, wie z.B. Arrhenatherum elatius (L.) P. Beauv. ex J. Presl & C. Presl, Plantago lanceolata und Achillea millefolium ssp. millefolium. Insgesamt ist die Entwicklung des ehemaligen Ackers jedoch positiv zu bewerten und eine weitere floristische Entwicklung in Richtung der Vegetation der Altheide ist zu erwarten.
Perspektiven für die zukünftige Entwicklung der „Garchinger Heide“
Eine Besonderheit des Naturschutzgebiets „Garchinger Heide“ ist das Nebeneinander von verschiedenen Standorten mit unterschiedlichen Bedingungen für das Pflanzenwachstum. Dadurch bietet die „Garchinger Heide“ Lebensraum sowohl für Arten der Halbtrockenrasen als auch der Trockenrasen und damit eine hohe Artenvielfalt. Diesen Zustand gilt es zu erhalten.
Im Bereich der Altheide kam es allerdings aufgrund des Stickstoffeintrags durch die Luft und des während der 1990er Jahre bis 2003 nur zweijährigen Mahdturnus stellenweise zu einer Vergrasung und Verfilzung der Vegetation, die einen Rückgang der lichtbedürftigen Magerrasen- und Trockenrasenarten bewirken kann. Um eine weitere Vergrasung und Streuakkumulation zu verhindern, ist eine Intensivierung des Mahdturnus der Altheide von zweijährig auf einjährig notwendig. Seit dem Jahr 2004 wird bei der Streifenmahd in der Altheide nur noch jeder fünfte Streifen (und nicht wie zuvor jeder zweite) als Rückzugsraum für Insekten stehen gelassen. Ob diese Mahdintensivierung ausreicht, bleibt zwar abzuwarten, auf dem ehemaligen Acker führte der bisherige einjährige Turnus jedoch bereits trotz der hohen Deckung der Süßgräser zu einer geringeren Deckung der Streu als in der Altheide. Eine Aushagerung durch jährliche Mahd ist auf dem ehemaligen Acker dennoch weiterhin notwendig, um die Etablierung der an nährstoffarme Böden angepassten Arten der Trocken- und Halbtrockenrasen zu fördern und die Ausbreitung potentiell dominanter Arten zu verhindern.

Abb. 13 Streifenmahd (August 2006); Foto: Daniela Röder
Fig. 13 Mowing in alterning stripes (August 2006); Photo: Daniela Röder
Um die Entwicklung des ehemaligen Ackers hin zur Altheide zu unterstützen, wäre es zudem von Vorteil, offene Bodenstellen zur Keimung und Etablierung von Magerrasenarten zu schaffen. Dies könnte alle zwei bis fünf Jahre im Anschluss an die Mahd durch maschinelles Rechen (z.B. mit Wiesenegge) geschehen, wodurch die hohe Moosdeckung vermindert würde.
Auch eine Wiedereinführung der Beweidung im Naturschutzgebiet könnte der Vergrasung und Verfilzung entgegen wirken (Hurst und John 1999, Jacquemyn et al. 2003). Auf dem ehemaligen Acker könnte durch Beweidung zudem die Einwanderung von Diasporen aus der Altheide durch zoochore Ausbreitung gefördert werden (vgl. Gibson und Brown 1991, Fischer et al. 1996). Durch die trittbedingte Schaffung von Regenerationsnischen können sich Zielarten möglicherweise besser etablieren, so dass sich die Vegetation des ehemaligen Ackers schneller der der Altheide angleicht.
Das Rollfeld als Sonderstandort und Lebensraum für seltene konkurrenzschwache Trockenrasenarten sollte als solcher erhalten bleiben, da die auf offene Standorte angewiesenen Trockenrasenarten sich nur über kurze Distanzen ausbreiten und neu geschaffene Abtragsflächen nur sehr langsam besiedeln (Will et al. 2005). Die gelegentliche Entbuschung des Rollfeldes sollte daher beibehalten werden, um der fortschreitenden Sukzession entgegenzuwirken. Durch gelegentliches Abrechen der zunehmend durch pleurokarpe Moose und Cladonien dominierten Moos- und Flechtenschicht könnten zudem nicht nur schwachwüchsige Gefäßpflanzen, sondern auch seltene akrokarpe Moose und Erdflechten gefördert werden (vgl. Günzl 2001, Jeschke und Kiehl 2006).
Nach Fischer (2003) ist die Schaffung neuer Abtragsflächen als Habitat für rohbodenbesiedelnde Pflanzen- und Tierarten zu empfehlen. Die Untersuchungen von Will et al. (2005) zeigen, dass sich auf einer 1980 angelegten Abtragsfläche, die sich nördlich an das Rollfeld anschließt, zahlreiche konkurrenzschwache Pflanzenarten der Trockenrasen ansiedeln konnten. Auf der südlich anschließenden Abtragsfläche von 2000 etablierten sich bis 2004 nur wenige Pflanzenindividuen, was zeigt wie langsam die Besiedlung neuer Flächen vonstatten geht.
Die Anlage weiterer Bodenabtragsflächen als Lebensraum für spezialisierte Rohbodenbesiedler sollte nicht im Naturschutzgebiet selbst erfolgen, in dem der Schutz und die Entwicklung der bestehenden Trocken- und Halbtrockenrasen mit Hilfe der oben genannten Pflegemaßnahmen Priorität hat. Sinnvoll ist der Oberbodenabtrag dagegen bei der Anlage neuer Magerrasen auf ehemaligen Äckern, die direkt an das Naturschutzgebiet angrenzen (s. Pfadenhauer und Kiehl 2003). Das Beispiel der 1993 angelegten Bodenabtragsfläche 506A zeigt, dass durch Bodenabtrag in Kombination mit Mähgutübertragung günstige Bedingungen für zahlreiche Zielarten des Naturschutzes geschaffen werden können (Jeschke und Kiehl 2006, Kiehl und Wagner 2006). Durch das Nebeneinander unterschiedlich alter Bodenabtragsflächen im Naturschutzgebiet selbst und auf den umliegenden Renaturierungsflächen können sowohl Pionierarten initialer Magerrasen (z.B. Biscutella laevigata, Leontodon incanus) als auch langsam wachsende konkurrenzschwache Chamaephyten, wie z.B. Globularia cordifolia oder Polygala chamaebuxus geeignete Lebensräume finden.
Literatur
Ad-Hoc-Arbeitsgruppe Boden (2005) Eckelmann, W. (Red.); Sponagel H, Grottenthaler W, Hartmann K-J, Hartwich R, Janetzko P, Joisten H, Kühn D, Sabel K-J, Traidl R: Bodenkundliche Kartieranleitung. 5. verbesserte und erweiterte Auflage. Bundesanstalt für Geowissenschaften und Rohstoffe in Zusammenarbeit mit den Staatlichen Geologischen Diensten der Bundesrepublik Deutschland (Hrsg.), Hannover. E. Schweizerbart’sche Verlagsbuchhandlung, Stuttgart
Bayerisches Landesamt für Umweltschutz (Hrsg.) (2000) Bestimmungsschlüssel für Flächen nach Art. 13d(1) BayNatSchG, Augsburg
Bender O, Boehmer HJ, Jens D, Schumacher KP (2005) Analysis of land-use change in a sector of Upper Franconia (Bavaria,Germany) since 1850 using land register records. Landscape Ecology 20: 149-163
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate – a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society. Series B 57: 289–300
Beinlich B, Manderbach D (1995) Die historische Landschafts- und Nutzungsentwicklung in Württemberg unter besonderer Berücksichtigung der Schwäbischen Alb. Beihefte zu den Veröffentlichungen für Naturschutz und Landschaftspflege in Baden-Würtemberg 83: 65-86
Bobbink R, Willems JH (1987) Increasing dominance of Brachypodium pinnatum (L.) Beauv. in chalk grasslands: A threat to a species-rich ecosystem. Biological Conservation 40: 301-314
Carroll JA, Caporn SJM, Johnson D, Morecroft M, Lee JA (2003) The interactions between plant growth, vegetation structure and soil processes in semi-natural acidic and calcareous grasslands receiving long-term inputs of simulated pollutant nitrogen deposition. Environmental Pollution 121: 363-376
Council of Europe (1979) Convention on the Conservation of European Wildlife and Natural Habitats. http://conventions.coe.int/Treaty/en/Treaties/Html/104.htm (30.08.2005)
Dengler J (2005) Zwischen Estland und Portugal - Gemeinsamkeiten und Unterschiede der Phytodiversitätsmuster europäischer Trockenrasen. Tuexenia 25: 387-405.
Dufrene M, Legendre P (1997) Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs 67: 345-366
During HJ, Willems JH (1984) Diversity models applied to a chalk grassland. Vegetatio 57: 103-114
Fetzer KD, Grottenthaler W, Hofmann B, Jerz H, Rückert G, Schmidt F, Wittmann O (1986) Standortkundliche Bodenkarte von Bayern 1:50 000 München-Augsburg und Umgebung. Erläuterungen. Bayerisches Geologisches Landesamt, München
Fischer S, Poschlod P, Beinlich B (1996) Experimental studies on the dispersal of plants and animals on sheep in calcareous grasslands. Journal of Applied Ecology 63: 1206-1221
Fischer FP (2003) Langzeitmonitoring von Heuschräkenbeständen im NSG „GarchingerHeide“ 1994-2001. In: Pfadenhauer J, Kiehl K (Hrsg.) Renaturierung von Kalkmagerrasen - Angewandte Landschaftsökologie 55: 201-209
Geisel O (1989) Von den Anfängen zum heutigen Naturschutzgebiet – und die Zukunft? In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 76-80
Gibson CWD, Brown VK (1991) The nature and rate of development of calcareous grasslands in southern Britain. Biological Conservation 58: 297-316
Günzl B (2001) Die Bunte-Erdflechten-Gesellschaft (Toninio-Psoretum decipientis Stodiek 1937) in Nordhessen - aktuelle Erfassung und Gliederung. Tuexenia 21: 179-191
Hepp E, Poelt J (1970) Die Garchinger Heide. Berichte der Bayerischen Botanischen Gesellschaft 42: 5-14
Hurst H, John E (1999) The biotic and abiotic changes associated with Brachypodium pinnatum dominance in chalk grassland in south-east England. Biological Conservation 88: 75-84
Hutchings MJ, Booth KD (1996) Studies on the feasibility of re-creating chalk grassland vegetation on ex-arable land. I. The potential roles of the seed bank and the seed rain. Journal of Applied Ecology 33: 1171-1181
Jacquemyn H, Brys R, Hermy M (2003) Short-term effects of different management regimes on the response of calcareous grassland vegetation to increased nitrogen. Biological Conservation 111: 137-147
Jeschke M, Kiehl K (2006) Auswirkung von Renaturierungs- und Pflegemaßnahmen auf die Artenzusammensetzung und Artendiversität von Gefäßpflanzen und Kryptogamen in neu angelegten Kalkmagerrasen. Tuexenia 26: 223-242
Kaufhold G (1990) Modellvorhaben Sicherung und Verbesserung der Heideflächen im Norden von München. Schriftenreihe Bayerisches Landesamt für Umweltschutz 105: 7-25
Kiehl K, Thormann A, Pfadenhauer J (2006) Evaluation of initial restoration measures during the restoration of calcareous grasslands on former arable fields. Restoration Ecology 14: 148-156
Kiehl K, Jeschke M (2005) Erfassung und Bewertung der Phytodiversität ursprünglicher und neu angelegter Kalkmagerrasen der nördlichen Münchner Schotterebene. Tuexenia 25: 445-461
Kiehl K, Wagner C (2006) Effects of hay transfer on long-term establishment of vegetation and grasshoppers on former arable fields. Restoration Ecology 14, 157-166
Kiehl K, Thormann A, Pfadenhauer J (2003) Nährstoffdynamik und Phytomasseproduktion in neu angelegten Kalkmagerrassen auf ehemaligen Ackerflächen. In: Pfadenhauer J, Kiehl K (Hrsg.) Renaturierung von Kalkmagerrasen - Angewandte Landschaftsökologie 55: 39-71
Kollmansberger G, Geisel O (1989a) Archäologen auf der Heide. In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 54-60
Kollmansberger G, Geisel O (1989b) Hochäcker in Heide und Lohe. In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 63-64
Kollmansberger G, Geisel O (1989c) Die Rettung der Garchinger Heide. In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 73-75
Kollmansberger G, Geisel O (1989d) Soldaten auf der Heide. In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 66-67
Korneck D, Müller T, Oberdorfer, E (1993) Sand- und Trockenrasen, Heide- Borstgras-Gesellschaften, alpine Magerrasen, Saum-Gesellschaften, Schlag- und Hochstauden-Fluren. In: Oberdorfer E. (Hrsg.) Süddeutsche Pflanzengesellschaften. Teil 2. Gustav Fischer Verlag, Jena
Lang A, Walentowski H, Lorenz W (2004) Kartieranleitung für die Lebensraumtypen nach Anhang I der Fauna-Flora-Habitat-Richtlinie in Bayern, 5. Entwurf. Bayer. Landesamt für Umweltschutz, Bayer. Landesanstalt f. Wald und Forstwirtschaft, Augsburg, Freising
Leuschner C (1989) Zur Rolle der Wasserverfügbarkeit und des Stickstoffangebots als limitierende Standortsfaktoren in verschiedenen basiphytischen Trockenrasen-Gesellschaften des Oberelsaß, Frankreich. Phytocoenologia 18: 1-54
McCune B, Mefford MJ (1999) Multivariate analysis of ecological data. Version 4.27, MjM Software, Gleneden Beach, Oregon, U.S.A.
Miller U, Pfadenhauer J (1997) Renaturierung von Kalkmagerrasen. Zur Vorhersage der gelenkten Sukzession durch Aufbringung von diasporenhaltigem Mähgut. Verhandlungen der Gesellschaft für Ökologie 27: 155-163
Neitzke M (1998) Changes in nitrogen supply along transects from farmland to calcareous grassland. Zeitschrift für Pflanzenernährung und Bodenkunde 161: 639-646
Oberdorfer E (2001) Pflanzensoziologische Exkursionsflora für Deutschland und angrenzende Gebiete. 8. Aufl., Eugen Ulmer Verlag, Stuttgart
Peet RK, Glenn-Lewin DC, Walker Wolf J (1983) Prediction of man's impact on plant diversity: a challenge for vegetation science. In: Holzner W., Werger MJ, Ikusima I (eds.) Man's impact on vegetation. Dr. W. Junk, The Hague: 41-54
Pegtel DM, Bakker JP, Verweij GL, Fresco LFM (1996) N, K, and P deficiency in chronosequential cut summer-dry grassland on gley podsol after the cessation of fertilizer application. Plant and Soil 178: 121-131
Pfadenhauer J (2002) Landnutzung und Biodiversität – Beispiele aus Mitteleuropa. Laufener Seminarbeitrag 2/02: 145-159
Pfadenhauer J (2001) Some remarks on the socio-cultural background of restoration ecology. Restoration Ecology 9: 220-229
Pfadenhauer J, Kiehl K (2003) Renaturierung von Kalkmagerrasen – Ein Überblick. In: Pfadenhauer J, Kiehl K (Hrsg.) Renaturierung von Kalkmagerrasen. Angewandte Landschaftsökologie 55: 25-38
Pfadenhauer J, Liebermann C (1986) Eine geobotanische Dauerbeobachtungsfläche im Naturschutzgebiet Garchinger Heide. Berichte der Bayerischen Botanischen Gesellschaft 57: 99-110
Pfadenhauer J, Miller U (2000) Verfahren zur Ansiedlung von Kalkmagerrasen auf Ackerflächen. In: Pfadenhauer J, Fischer FP, Helfer W, Joas C, Lösch R, Miller U, Miltz C, Schmid H, Sieren E, Wiesinger K Sicherung und Entwicklung der Heiden im Norden von München.. Angewandte Landschaftsökologie 32: 37-87
Poschlod P, WallisDeVries MF (2002) The historical and socio-economic perspective of calcareous grasslands. Lessons from the distant and recent past. Biological Conservation 104: 361-376
Quinger B, Bräu M, Kornprobst M (1994) Lebensraumtyp Kalkmagerrasen. Landschaftspflegekonzept Bayern - Band II.1. Bayerisches Staatsministerium für Landesentwicklung und Umweltfragen & Bayerische Akademie für Landschaftspflege und Naturschutz, München
Rennwald E (2000) Verzeichnis und Rote Liste der Pflanzengesellschaften Deutschlands. Schriftenreihe für Vegetationskunde 35
Riemenschneider M (1956) Vergleichende Vegetationsstudien über Heidewiesen im Isarbereich. Berichte der Bayerischen Botanischen Gesellschaft 31: 75-120
Ringler A (2002) Magerrasen in einer neuorientierten Agrarlandschaft. Schriftenreihe des Bayerischen Landesamts für Umweltschutz 167: 9-22
Röder D, Kiehl K (2006) Population structure and population dynamic of Pulsatilla patens (L.) MILL. in relation to vegetation characteristics. Flora 201 (in Druck)
Schauer T (1969) Die Flechtenvegetation der Kiesflache auf der Garchinger Heide nordlich von München. Herzogia 1: 181-186
Scheffer F, Schachtschabel P (2002) Lehrbuch der Bodenkunde. 15. Auflage. Spektrum Akademischer Verlag, Heidelberg
Scheuerer M, Ahlmer W (2003) Rote Liste gefährdeter Gefäßpflanzen Bayerns mit regionalisierter Florenliste. Schriftenreihe Bayerisches Landesamt für Umweltschutz 165
Schlichting E, Blume H-P, Stahr K (1995) Bodenkundliches Praktikum: eine Einführung in pedologisches Arbeiten für Ökologen, insbesondere Land- und Forstwirte und für Geowissenschaftler. 2. Auflage. Blackwell Wissenschaftsverlag, Berlin
Sendtner O (1854) Vegetations-Verhältnisse Südbayerns nach den Grundsätzen der Pflanzengeographie und mit Bezugnahme auf Landescultur. Literarisch-artistische Anstalt, München
Skinner RJ, Todd AD (1998) Twenty-five years of monitoring pH and nutrient status of soils in England and Wales. Soil Use and Management 14: 162-169
Unkovich M, Jamieson N, Monaghan R, Barraclough D (1998) Nitrogen mineralization and plant nitrogen acquisition in a nitrogen-limited calcareous grassland. Environmental and Experimental Botany 40: 209-219
VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten) (1991) Methodenbuch Band I.: Die Untersuchung von Böden. A 5.1.1. VDLUFA-Verlag, Darmstadt
VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten) (2002) Methodenbuch Band I.: Die Untersuchung von Böden. 3. Teillieferung A 6.2.1.1. VDLUFA-Verlag, Darmstadt
Verhoeven KJF, Simonsen KL, McIntyre LM (2005) Implementing false discovery rate control: increasing your power. Oikos 108: 643-647
Vollmann F (1911) Das Schutzgebiet der Bayerischen Botanischen Gesellschaft auf der Garchinger Heide. Mitteilungen der Bayerischen Botanischen Gesellschaft 2: 312-318
Wells TCE, Sheail J, Ball DF, Ward LK (1976) Ecological studies on the Porton Ranges: Relationships between vegetation, soils and land-use history. Journal of Ecology 64: 589-626
Will H, Eichinger N, Röder D, Kiehl K (2005) Vergleich der Vegetation unterschiedlich alter Bodenabtragsflächen im Naturschutzgebiet Garchinger Heide. Berichte der Bayerischen Botanischen Gesellschaft 75: 169-180
Willems JH, Peet RK, Bik L (1993) Changes in chalk-grassland structure and species richness resulting from selective nutrient additions. Journal of Vegetation Science 4: 203-212
Willems JH (2001) Problems approaches and results in restoration of Dutch calcareous grasslands during the last 30 years. Restoration Ecology 9: 147-154
Windolf R (1989) Die geologische Geschichte des G`filds. In: Gemeinde Eching, Landkreis Freising (Hrsg.) Garchinger Heide, Echinger Lohe, Naturschutzgebiete in der Gemeinde Eching, Landkreis Freising. Bruckmann Verlag und Druck GmbH, München: 13-15
Wittmann O (1983) Standörtliche Landschaftsgliederung von Bayern. Bayerisches Geologisches Landesamt, München
Anhang / Appendix
1
Deckung aller gefundenen Gefäßpflanzen (in %) sortiert nach Artengruppen, die nur auf dem Rollfeld, in der Altheide oder auf dem ehemaligen Acker vorkommen bzw. dort eine höhere Stetigkeit ( jeweils mind. 10%-Punkte mehr als in den anderen Gebieten) und eine höhere Deckung als auf den anderen Standorten haben.
Cover of all Phanerogams found (in %) arranged by groups of species which occurred only on the "Rollfeld" , the ancient grassland and the ex-arable field, or which occurred on these areas with higher frequency (> 10 %) or higher cover (> 10 %) than on the other sites.
Microsoft Excel (180 KB)
Adobe PDF (20 KB)
2
Abkürzungen der Arten der DCA (siehe Abb. 9)
abbreviations of species names in the DCA (see Fig. 9)
achmil: Achillea millefolium
adover: Adonis vernalis
allcar: Allium carinatum
antram: Anthericum ramosum
antvul: Anthyllis vulneraria
arrela: Arrhenatherum elatius
aspcyn: Asperula cynanchica
asptin: Asperula tinctoria
avepra: Helictotrichon pratensis
betoff: Betonica officinalis
bislae: Biscutella laevigata
brapin: Brachypodium rupestre
brimed: Briza media
broere: Bromus erectus
bupsal: Buphthalmum salicifolium
camglo: Campanula glomerata
camrot: Campanula rotundifolia
caraca: Carlina acaulis
carcar: Carex caryophyllea
cardef: Carduus defloratus
careri: Carex ericetorum
carhum: Carex humilis
carmon: Carex montana
cenjac: Centaurea jacea ssp. angustifolia
censca: Centaurea scabiosa
charat: Chamaecytisus ratisbonensis
corvag: Coronilla vaginalis
dansp: Danthonia decumbens
dapcne: Daphne cneorum
dorger: Dorycnium germanicum
eriher: Erica herbacea
eupcyp: Euphorbia cyparissias
eupver: Euphorbia verrucosa
fesrub: Festuca rubra
fesrup: Festuca rupicola
filvul: Filipendula vulgaris
galbor: Galium boreale
galmol: Galium mollugo
genclu: Gentiana clusii
gentin: Genista tinctoria
genver: Gentiana verna
glocor: Globularia cordifolia
glopun: Globularia punctata
gymcon: Gymnadenia conopsea
helnum: Helianthemum nummularium ssp. obscurum
hiepia: Hieracium pilosella
hipcom: Hippocrepis comosa
inuhir: Inula hirta
koepyr: Koeleria pyramidata
leohis: Leontodon hispidus
leoinc: Leontodon incanus
lincat: Linum catharticum
linper: Linum perenne
lotcoc: Lotus corniculatus ssp. corniculatus
lotcoh: Lotus corniculatus ssp. hirsutus
peuore: Peucedanum oreoselinum
pimsax: Pimpinella saxifraga
plalan: Plantago lanceolata
plamed: Plantago media
polama: Polygala amarella
polcha: Polygala chamaebuxus
polodo: Polygonatum odoratum
polvul: Polygala vulgaris
potalb: Potentilla alba
potare: Potentilla incana
prugra: Prunella grandiflora
pulpat: Pulsatilla patens
pulvul: Pulsatilla vulgaris
rhasax: Rhamnus saxatilis
rhigla: Rhinanthus glacialis
salpra: Salvia pratensis
salele: Salix eleagnos
sanmin: Sanguisorba minor
scacan: Scabiosa canescens
scacol: Scabiosa columbaria
senjac: Senecio jacobaea
silnut: Silene nutans
taroff: Taraxacum officinale agg.
teumon: Teucrium montanum
thelin: Thesium linophyllon
thypra: Thymus praecox
trimon: Trifolium montanum
verspi: Veronica spicata
viohir: Viola hirta